Integrating Biodiversity into Land Use Planning
 |
 |
 |
 |
|||||
Session:Biodiversity Information for Local Planners (March 12, 10:15am)
|
|||||||||||||||||||||||||||
|
Integrating Biodiversity into Land Use Planning
|
|
||||||||||||||||||||||||||
Session:Biodiversity Information for Local Planners (March 12, 10:15am) |
|||||||||||||||||||||||||||
|
Abstract: Preservation of biodiversity is one of the most contentious environmental issues in the United States. Researchers have made progress in improving our understanding of the impacts of development but this has not led to much improvement in planning tools. This talk will offer a conceptual framework for addressing biodiversity-related planning questions. A model for selecting new biological reserves will be described, both to illustrate some of the lessons learned and future directions needed to incorporate biodiversity management into the larger arena of land use planning. The key to progress is better integration among scientists from different disciplines.
IntroductionCompetition for land among proponents of the built environment, working landscapes (farm and rangeland and resource extraction areas), and natural environments is one of the most contentious issues in the United States. Land use change, including urbanization, that consumes native habitat is believed the primary factor in loss of biodiversity (Foin et al. 1998, Wilcove et al 1998). Not only does urban expansion, and other land uses, consume habitat, but it also divides habitat into smaller fragments that may become too small and isolated to support viable populations for some species. Population growth, market economic forces, and private property rights continually apply pressure to convert an ever-increasing proportion of our finite land base into uses that are less compatible with maintaining the complete ensemble of biodiversity. Against this onslaught, conservation faces a formidable challenge. Most of our actions are taken as a last resort, i.e., once species have already become endangered and therefore covered by federal and state legislation. Another common approach is a "skirmish" strategy, employing legal action to block proposed development projects, often after they are far along in the decision process. This is costly to all stakeholders and fosters increasing rancor between the parties. A final strategy is to buy a threatened property and establish a nature reserve. These usually require a willing seller and are often purchased at huge expense to taxpayers or to conservation organizations (e.g., the "Headwaters" redwood forest in California). Selecting sites for new reserves is often opportunistic and seldom done with a systematic methodology. Acting at the 11th hour is costly and precludes any flexibility in how conservation goals might alternatively be achieved. It is difficult with these "ad hoc" approaches to determine the relative importance of a site–a case can be made that every parcel has some conservation value. Planners are caught in the middle of these competing social values. I suggest that conservation of biological diversity will be more cost-effective and socially acceptable when it is integrated into the land use planning process more explicitly than it currently is. Planners have the challenge, but I would argue also the opportunity, to do so. Given that there is interest in integrating biodiversity considerations into land use planning, what are the obstacles that restrict this integration? Perhaps the major obstacle is that the term "biodiversity" is a shorthand notation for a whole suite of complex biological and ecological features and processes that are hard to think about operationally. It is like "freedom." Everyone is for it but what do we actually do about it? How much can we afford to lose? And when is the price too dear? Without an operational set of variables, naturally it is hard for biodiversity to compete with social values that are easily measured or forecast such as jobs, tax revenues, or per capita income. Even where we attempt to quantify key aspects of biodiversity, and can find appropriate data for them, we still have difficulty relating policy and planning decisions to impacts on biodiversity. Of the many possible questions about biodiversity that planners might want to address, the following are particularly relevant:
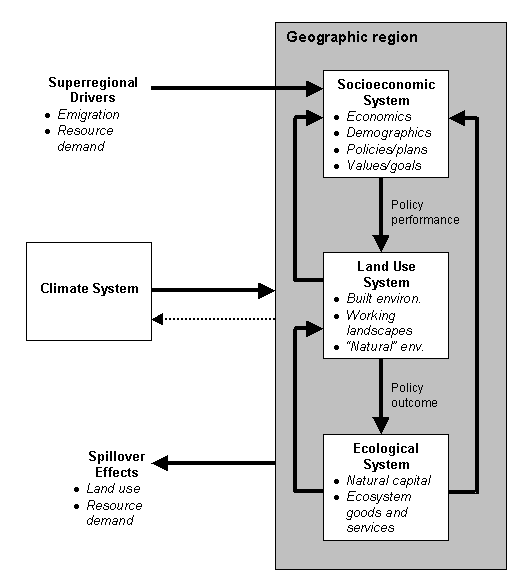
The objectives of this paper are to offer a framework that conceptually shows how these questions can be integrated into the planning process, illustrate one particular area where research has been transferred into planning practice, and use that to sketch areas where future research is still required. A Conceptual FrameworkCertainly part of the barrier to integration of biodiversity considerations into land use planning is a lack of communication between academic scientists and practicing planners (Prendergast et al. 1999). The good news is that science is making headway that planners should know about. It is useful to develop a conceptual framework of the pieces of the planning puzzle and the processes of their interaction. With such a framework, I hope to illustrate both the progress being made toward methods to answer the questions listed above and the remaining obstacles. These interrelationships can be visualized many ways, but I propose the following conceptual framework (Figure 1). The boxes represent systems, and the arrows are dynamic processes. In this framework, a geographic region contains three systems–Socioeconomic, Land Use, and Ecological. In addition, there are external drivers shaping the internal systems and over which the local decision makers have little control. For instance, national and international demand for natural resources (or for conservation) influence local decisions but local policy can do little to change the demand itself. Changes in the Socioeconomic System, moderated by constraints from the Climate and Ecological Systems, spur changes in the Land Use System. Land use changes then have impacts on the "natural capital" and ecosystem goods and services of the Ecological System, which responds indirectly to policies and plans. Thus Questions #1 and #3 are not so straightforward without modeling future land use change first. Policy interventions, such as siting new reserves (Question #4), can be applied in the Socioeconomic System, but their performances are measured in the effects on the Land Use System. The outcomes of those interventions are measured by changes (or prevention of negative effects) in the Ecological System (Oñate et al. 2000). Assessment of the Ecological System can address Question #2 to rank conservation values, but this information must be fed into the Socioeconomic System to compete with other values and goals. Questions #5-7 are by far the most complex, because they involve interaction among all three systems over time as well as the answers to the other questions. I include "spillover effects" in the framework as a reminder that decisions made locally often have implications on neighboring geographic regions. For instance, a decision to save habitat may simply deflect the demand for more housing somewhere else.
Progress has been made on some of these planning questions. Researchers have developed methods that predict impacts on species and habitats from simulations of urban growth (Question #1) (White et al. 1997), Landis et al. 1998), Duane 1999), and Cogan in press). Perhaps the greatest progress has been made in translating conservation principles into computerized strategies for exploring alternative networks of biodiversity management areas or reserves (Question #4). Questions #5-7 have scarcely been addressed. In the next sections I will briefly sketch the general problem of reserve selection and different approaches and then illustrate it with a specific spatial decision support tool. This planning tool exemplifies both the progress made in conservation planning and remaining issues that are being addressed in ongoing research. General Formulation of the Reserve selection problemAlthough conservation planning may entail many complementary approaches to protection of biological diversity, selecting a network of core reserves dedicated primarily to conservation is fundamental to most strategies. Reserve system design is a difficult planning problem because the number of species or ecosystems and locations can be quite large and beyond the capacity of an analyst to solve unaided. Over the past twenty years, conservation planners have developed increasingly sophisticated computer-based approaches to make the reserve site selection process more systematic and explicit. Reserve site selection entails identifying a set of locations that collectively capture viable examples of all native species and plant communities from among a set of "planning units" within a planning region. Most models begin with data on biodiversity elements (species, plant communities, and biophysical environments) at locations or planning units. The data may be in the form of {0,1} for absence or presence of the element, a probability of occurrence, or the extent of the element in the planning unit. The objective to be solved in reserve selection models is either to select the minimum number (or area) of planning units to contain all elements at a prescribed level (set covering) OR to contain the most elements at some fixed cost (maximum covering). Early models followed simple heuristic sets of rules to select reserve sites (Kirkpatrick 1983). More recent models adapted algorithms from operations research to identify optimal solutions (Church et al. 1996, 2000). These models generally respond to the perceived need for reserve siting to be as efficient or cost-effective as possible, given the competing social and economic demands for land and water. Variations of the models allow for weighting of species (such as by their rarity, Gerrard et al. 1997) or of sites (by the quality of habitat, Church et al. 2000). Let’s examine one reserve selection model to show what they do and don’t do. Sites modelSites was developed at the University of California, Santa Barbara, to meet the needs of The Nature Conservancy’s ecoregional planning process. Sites is a customized ArcView project that facilitates designing and analyzing alternative conservation portfolios (Andelman et al. 1999). The software in Sites to select of regionally representative systems of nature reserves is called the Site Selection Module (SSM), a streamlined derivative of SPEXAN (Possingham et al. 2000). I use Sites here not because it is the best model, but because it is one of the few models that has made the transition from research to operational planning tools. The overall objective of the reserve selection process in Sites is to minimize the "cost" of the reserve network while attempting to meet all conservation goals. Representation goals could be the specified number of population occurrences of a species or total area of each plant community type that must be included in the regional network. Spatial configuration goals specify the relative importance of selected sites being contiguous to achieve spatial compactness and connectivity. The SSM represents this set of objectives as: Total Cost = (cost of selected sites) + (penalty cost for not meeting the stated conservation goals for each element) + (cost of spatial dispersion of the selected sites as measured by the total perimeter of the set of reserves). The algorithm thus seeks to minimize Total Cost by selecting that set of planning units that covers as many elements as possible as cheaply as possible in as compact a set of sites as possible. It is also possible to specify a budget constraint, which limits the actual cost but incurs a greater penalty cost for the conservation targets that cannot be met. Sites can be used to explore alternative reserve systems in which planners can vary the budget, the importance of clustering reserves, the conservation goals, the importance of individual conservation elements, and so on. Since it is integrated into a GIS environment, it is simple to select planning units that either must be in the network or are precluded from selection. Thus planners could begin with a suite of alternatives representing different stakeholder views, lock in places where there is consensus, and iterate within the smaller decision space. During these iterations, local knowledge can be used to rule out unsuitable places (e.g., disturbed habitat, unwilling seller). GIS functions can be used to evaluate the impacts of an alternative reserve network on other resource and socioeconomic values. Even if the final reserve plan does not formally employ Sites, or a similar computer model, the exercise is helpful in making the goals explicit, in evaluating and exploring the decision space, and in answering some of the questions we posed at the beginning of this paper. Current Limitations and future directionsIt is generally assumed in reserve selection models that meeting conservation goals for specific amounts of habitat for each species or other biodiversity surrogate will achieve the ultimate goal of maintaining viable populations of native species. This assumption is almost never tested, in part because reserve selection models only deal with changes in the Land Use System of the conceptual framework (Figure 1) and do not assess the impacts in the Ecological System. These models ignore land uses and habitat patterns outside of reserves. Reserve selection models also do not consider the interaction between fragmented habitat patches, which can significantly affect viability through rates of colonization and gene flow. Testing viability and extinction risk of populations can be done with metapopulation or population viability analysis models. These models could provide feedback, or at least rules-of-thumb, to reserve selection models about the spatial configuration of reserves (Pelletier 2000). Although conservation biologists use such models, they have not yet been integrated with reserve selection models. At most they have been used to assess impacts of alternative land use futures. It is increasingly recognized that the cost of acquiring reserves (and the associated opportunity costs of foregone resources) means that it will be unrealistic to rely exclusively on that strategy to meet conservation goals. Thus the conservation planning problem should ultimately expand to consider a range of management options and focus on reserves where that is the most cost-effective conservation strategy. We need models that explore policy options or market-incentives for conservation that explicitly determine tradeoffs between competing uses and where multiple objectives can be satisfied simultaneously. Despite a paradigm shift in ecology that now views ecosystems as dynamic interactions, reserve siting methods have generally assumed a static view of the landscape. We treat biodiversity as a collection of things and reserves as places to keep those things permanently. This perilously simplifies the complex reality of ecosystems. First it ignores the processes that are constantly influencing the composition, structure, and function, and therefore the biodiversity, of a landscape. This simplification can also mislead us into thinking that a tract of land can always be added to the reserve system at a future date so long as the land is not developed first. Ideally, future conservation planning models should account for the temporal dynamics of ecosystems, perhaps as probabilities of change, and should explicitly deal with the economic reality that reserves will be acquired over time as budgets allow. Considering how the probability of development, or other ecosystem change, varies across the landscape and over time may alter the prioritization of sites for new reserves. Climate change is the monster looming over the horizon that may finally force us to consider long-term changes in the location and content of ecosystems we are trying to protect. Land use simulation modeling is needed to predict the likely condition of the matrix and therefore its effect on reserves. Reserve siting models focus on meeting conservation goals, but are generally weak in their socioeconomic considerations. They typically attempt to be cost-effective by minimizing the cost (usually estimated by land area) of achieving the conservation goals. Data on real land values are seldom available. Even more rare is to see opportunity costs of foregone resources considered (except see Faith et al. 1996 for a forestry example). Conservation planning is fraught with uncertainties. Within the Ecological System, we generally do not know absolutely which species occur in sites, especially over time. Often we must infer their presence from simple surrogates such as suitable habitat. Reserve selection models typically ignore this uncertainty and treat the ecological data as facts. Only occasionally do researchers test models in which the species data are treated as probabilities rather than certainties (Polasky et al. 2000). Similar uncertainties abound in the Socioeconomic System as well. We estimate land values (and usually assume they are constant over time), and we assume that selected properties are currently owned by willing sellers. We also assume that unselected properties will be fully developed in the short-term. In other words, we model individualistic personal behaviors and decision making in the simplest, most generalized manner. For conservation planning research to reach a stage where practicing planners can really benefit from it, these various forms of uncertainty need to be explicitly modeled. Then planners can test the robustness of alternative plans to the assumptions and inherent uncertainties of the problem. Most of the pieces for integrating biodiversity into land use planning currently exist, at least as research tools. There are many land use change models that simulate the changes in the Land Use System triggered by factors in the Socioeconomic System. Reserve selection models have advanced in sophistication, particularly by adapting methods from operations research. Models exist to assess the risk of extinction of species populations in different geometric arrangements of habitat patches. Uncertainty is sometimes handled explicitly in each of these models, either as probabilities or sensitivity analysis. The current limitation is the lack of integration of these pieces. This is partly due to the legacy of a reductionist scientific culture that isolates specialists in their limited domains. Synthesis has been rare. Planning, in contrast, is by nature a very integrative activity. I believe, however, that we are on the verge of some major advances in the planning process because of the rising interaction of academic biologists, geographers, economists, and other disciplines in framing the conservation problem in more than just the ecological dimension. We seek to develop a new, multidisciplinary research program that integrates the modeling process for urban growth and biodiversity conservation. This program could provide valuable tools to planners and local decision-makers that account for biodiversity while accommodating growth. These tools will also evaluate alternative growth policies to determine their likely effectiveness in achieving biodiversity goals. As almost every metropolitan area in the country faces this dilemma, this multidisciplinary research program could have wide practical impact. ReferencesAndelman, S., I. Ball, F. Davis, and D. Stoms. 1999. Sites version 1.0. An Analytical Toolbox for Designing Ecoregional Conservation Portfolios. Manual prepared for The Nature Conservancy, December 1999. University of California, Santa Barbara, 55 pp. Church, R., R. Gerrard, A. Hollander and D. Stoms. 2000. Understanding the tradeoffs between site quality and species presence in reserve site selection. Forest Science 46: 157-167. Church, R. L., D. M. Stoms and F. W. Davis. 1996. Reserve selection as a maximal covering location problem. Biological Conservation 76: 105-112. Cogan, C. B. In press. Biodiversity conflict analysis at multiple spatial scales. In Predicting Species Occurrences: Issues of Accuracy and Scale. J.M. Scott, P. Heglund, M. Morrison, B. Wall, F. Samson, and J. Haufler, eds. Island Press, Covello, CA. Duane, T. P. 1999. Shaping the Sierra: Nature, Culture, and Conflict in the Changing West. University of California Press, Berkeley. 595 pp. Faith, D. P., P. A. Walker, J. R. Ive and L. Belbin. 1996. Integrating conservation and forestry production: Exploring trade-offs between biodiversity and production in regional land-use assessment. Forest Ecology and Management 85: 251-260. Foin, T. C., S. P. D. Riley, A. L. Pawley, D. R. Ayres, T. M. Carlsen, R. J. Hodum and P. V. Switzer. 1998. Improving recovery planning for threatened and endangered species. BioScience 48: 177-184. Gerrard, R. A., R. L. Church, D. M. Stoms, and F. W. Davis. 1997. Selecting conservation reserves using species covering models: Adapting the ARC/INFO GIS. Transactions in GIS 2: 45-60. Kirkpatrick, J. B. 1983. An iterative method for establishing priorities for selection of nature reserves: an example from Tasmania. Biological Conservation 25: 127-134. Landis, J. D., J. P. Monzon, M. Reilly and C. Cogan. 1998. Development and pilot application of the California Urban and Biodiversity Analysis (CURBA) model. Proceedings of 18th Annual ESRI User Conference, San Diego, ESRI. Published online at http://www.esri.com/library/userconf/proc98/PROCEED/TO600/PAP571/P571.HTM . Oñate, J. J., E. Andersen, B. Peco and J. Primdahl. 2000. Agri-environmental schemes and the European agricultural landscapes: the role of indicators as valuing tools for evaluation. Landscape Ecology 15: 271-280. Pelletier, J. D. 2000. Model assessments of the optimal design of nature reserves for maximizing species longevity. Journal of Theoretical Biology 202: 25-32. Polasky, S., J. D. Camm, A. R. Solow, B. Csuti, D. White and R. Ding. 2000. Choosing reserve networks with incomplete species information. Biological Conservation 94: 1-10. Possingham, H., I. Ball, and S. Andelman. 2000. Mathematical methods for identifying representative reserve networks. Pages 291-305 in: Quantitative methods for conservation biology. Ferson, S. and Burgman, M. (eds). Springer-Verlag, New York. Prendergast, J. R., R. M. Quinn and J. H. Lawton. 1999. The gaps between theory and practice in selecting nature reserves. Conservation Biology 13: 484-492. White, D., P. G. Minotti, M. J. Barczak, J. C. Sifneos, K. E. Freemark, M. V. Santelmann, C. F. Steinitz, A. R. Kiester and E. M. Preston. 1997. Assessing risks to biodiversity from future landscape change. Conservation Biology 11: 349-360. Wilcove, D. S., D. Rothstein, J. Dubrow, A. Phillips and E. Losos. 1998. Quantifying threats to imperiled species in the United States. BioScience 48: 607-615. AcknowledgmentsThe author was partially supported by a grant from the Research Across Disciplines program at the University of California, Santa Barbara. I appreciate the ideas and suggestions of Sandy Andelman, Antonio Bento, Chris Cogan, Chris Costello, Rick Church, Frank Davis, Doug Fischer, John Gallo, Greg Greenwood, Bruce Kendall, and others that helped shape this paper. The Sites 1.0 conservation planning software and users manual are available online at http://www.biogeog.ucsb.edu/projects/tnc/toolbox.html.
Author and Copyright InformationThe author is a research geographer and manager of the Biogeography Lab at the University of California, Santa Barbara. His research covers a range of topics in biodiversity assessment and conservation planning. He is currently a member of a working group at the National Center for Ecological Analysis and Synthesis in Santa Barbara that is developing a statewide conservation strategy for California’s Resources Agency. He has also been a planner for the U. S. Forest Service at Lake Tahoe, California. He can be contacted at stoms@icess.ucsb.edu. |